Fjær, lim og rakettbatterier i celledeling
Av David Coppedge, 11. juli 2024. Oversatt herfra
Når jeg nylig skrev her -lenke, sammenlignet jeg en celle som gjennomgikk mitose med en ranch der cowboyer vokter kromosomer i separate korraler. Hvis forskningen bak den artikkelen ikke var nok til å inspirere ærefrykt, er det mye mer. Her er sammendrag av andre forskningsartikler om celledeling.
Feilkorreksjon og sjekkpunkter
En artikkel i PNAs -lenke tar opp spørsmålet om hvordan spindelen fester seg til kromosomene og utfører "feilretting." Spindelen er en eike-lignende struktur som vokser ut fra sentriolene i sentrosomet. Spindelfibre, bestående av mikrotubuler (lange kjeder av tubulinproteiner som vokser og krymper dynamisk), vokser mot kromosomene, som er stilt opp i par langs den sentrale aksen (dette etter alt DNA har blitt duplisert og supercoiled i den kjente x -formen av kromosomer). Hver kromatid (en på hver side, forbundet med en centromer) må feste seg til en og bare en spindelmikrotubul. Men de starter ikke på den måten; Én kromatid kan ha flere, og andre har ingen. I artikkelen har Ha et al. lurt på hvordan de ender opp helt riktig. Hvordan fikses tilknytnings-feil?
Bilde 1. Centromer -sentral ved kromosom segregering
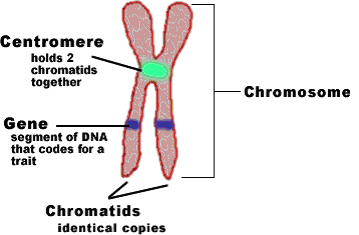 "Den mitotiske spindelen er en bipolar struktur, først og fremst sammensatt av mikrotubuler, som segregerer et like antall kromosomer til hver dattercelle med bemerkelsesverdig troskap. Spindelen starter med mange uriktige tilknytninger mellom kromosomer og mikrotubuler, som blir korrigert over tid for til slutt å tilfredsstille spindelenhetens sjekkpunkt, og lar cellen fortsette å segregere kromosomene. Kromosom missegregering tilskrives ofte feil i feilkorreksjon. Til tross for den utbredte viktigheten av kromosomsegregeringsfeil, er det foreløpig mangel på kvantitative metoder for å karakterisere dynamikken i feilkorreksjonsprosessen og hvordan det kan gå galt.
"Den mitotiske spindelen er en bipolar struktur, først og fremst sammensatt av mikrotubuler, som segregerer et like antall kromosomer til hver dattercelle med bemerkelsesverdig troskap. Spindelen starter med mange uriktige tilknytninger mellom kromosomer og mikrotubuler, som blir korrigert over tid for til slutt å tilfredsstille spindelenhetens sjekkpunkt, og lar cellen fortsette å segregere kromosomene. Kromosom missegregering tilskrives ofte feil i feilkorreksjon. Til tross for den utbredte viktigheten av kromosomsegregeringsfeil, er det foreløpig mangel på kvantitative metoder for å karakterisere dynamikken i feilkorreksjonsprosessen og hvordan det kan gå galt.
Et sjekkpunkt! Hvilket konsept! Cellen har mange GO/No-Go-sjekkpunkter som ikke lar celledelingen fortsette før feil er fikset. Dette høres ut som automatisert kvalitetskontroll av noen som hadde framsyn for å vite “hvordan det kan gå galt” og konstruert “feilretting” for å forhindre feil. Hvordan kan en blind darwinistisk prosesser noen gang komme fram til sjekkpunkter?
Forfatterne lærte at "feilkorreksjon er en kromosom-autonom prosess som inntreffer med konstant hastighet under spindelmontering." Sikkert, men andre prosesser ser på og overvåker, fordi sjekkpunkter og feilkorreksjon trosser darwinistiske forklaringer -lenke.
Når vi snakker om sjekkpunkter, kunngjorde forskere ved University of Duisburg-Essen identifiseringen av et av proteinene som gir 'altklart' signalet for mitose (celledeling). Standardtilstanden er no-go, eller stopp. "De oppdaget hvordan initiativtakeren til stoppsignalet, en proteinkinase kalt MPS1, er bundet til tilknytningsstedet til kromosomene og hvordan det bare løsner når kromosomene er riktig bundet til den mitotiske spindelen." Funnene ble detaljert i en artikkel i Current Biology -lenke.
Fjær og flaskebørster i centromerer
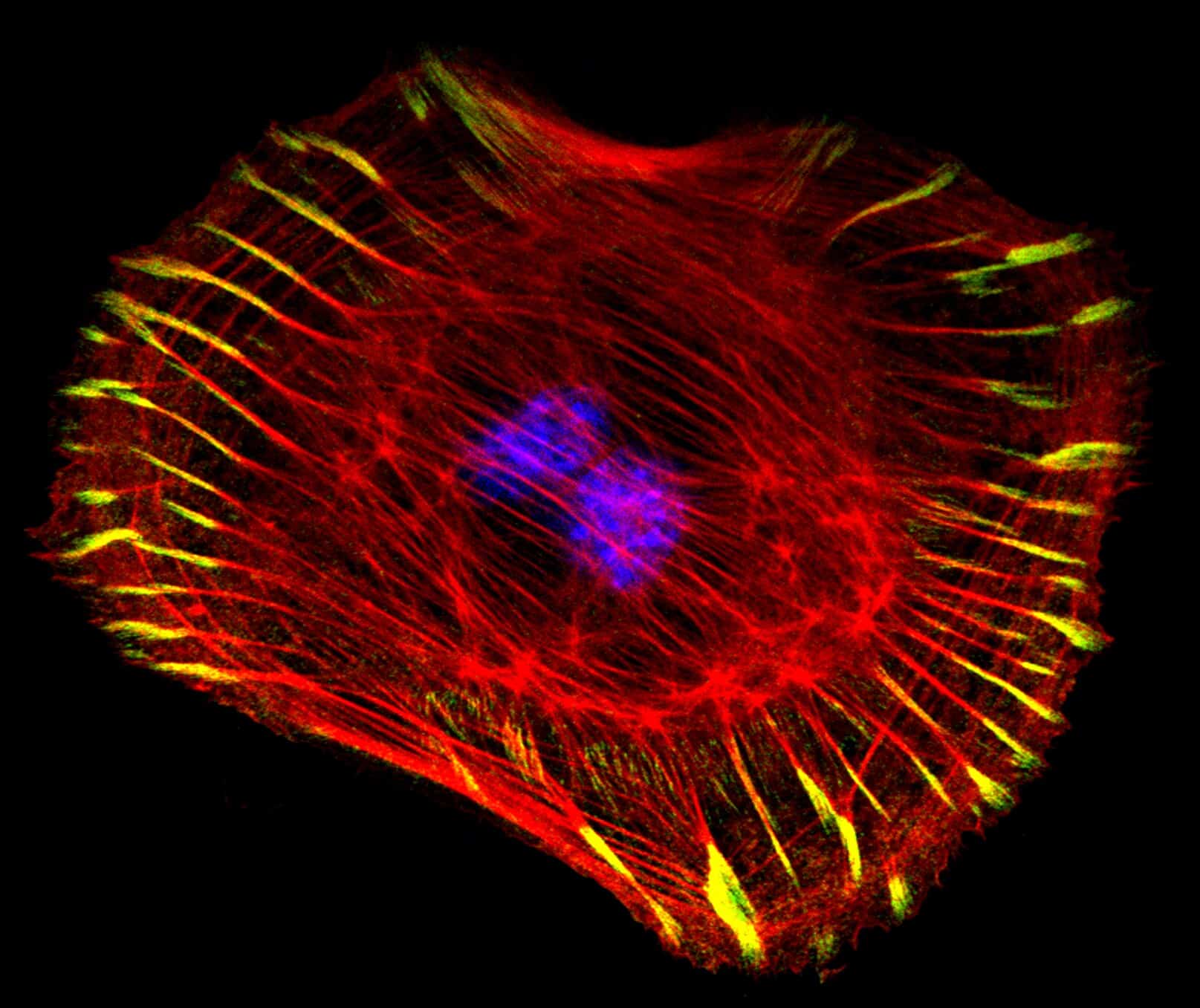 En utsendelse av Kerry Bloom in Current Biology -lenke, rapporterte om arbeid som viser at "åkene" i min analogi (centromerene) har en fjær-likhet ved seg, som hjelper til med å skape riktig spenning på tauene før "cowboyer" -vinsjet dem fra hverandre. Denne artikkelen understreker rollen til proteiner som fysiske strukturer, som er i stand til å formidle krefter til andre molekyler.
En utsendelse av Kerry Bloom in Current Biology -lenke, rapporterte om arbeid som viser at "åkene" i min analogi (centromerene) har en fjær-likhet ved seg, som hjelper til med å skape riktig spenning på tauene før "cowboyer" -vinsjet dem fra hverandre. Denne artikkelen understreker rollen til proteiner som fysiske strukturer, som er i stand til å formidle krefter til andre molekyler.
"Å vende centromert-DNA til en mekanisk fjær er sentral i troverdigheten til kromosom-segregering. En fersk studie viser hvordan centromere DNA -løkker og partisjonering av kohesin og kondensin, konverterer centromerer og peri-centromerer til todelte flaskebørster."
Bloom viser et diagram over "flaskebørste" -formen til disse centromer-proteinene. Formen, bestående av avstandsløkker i proteinet, gir fleksibilitet på centromeren, og holder den stiv, men fjærende nok til å henge på DNA-tråder.
Bilde 2. Celledeling pågår -celle strekkes ut til todelt 'flaskebørste'
"Hvert av centromerdomenene danner det som er kjent som en flaskebørste. En flaskebørste (Bilde 2) er fysikerens løsning for å transformere en diskettpolymer til en mekanisk fjær. En trengsel av sidekjeder (DNA -løkker) fra en primær akse reduserer svingninger langs aksen og genererer strekkfasthet. Når den primære aksen bøyer seg, reduseres avstanden mellom sidekjedene i retning av krumningen, noe som øker trengsel mellom sidekjedene som igjen motvirker krumningen"
Se her for mer om kohesin og kondensin, to essensielle proteiner involvert i DNA -pakking -lenke, "Et av de ypperste naturens underverk."
Artikkelen nevner en potensiell funksjon for ikke -kodende DNA i centromere: pakkematerialet! I det han tar opp spørsmålet om hvorfor centromerer er forskjellige i størrelse, skriver han:
"Sentromerer er også gåtefulle fra sekvensperspektivet. De kan være så små som 117 bp (spirende gjær), og opp til flere megabaser hos mennesker. De kan bygges på unike sekvenser eller hierarkiske matriser av små satellittrepetisjoner (171 bp alfa-satellitt). Dette paradokset kan løses ved å skille mellom plattformen for kinetokore montering og mikrotubule-binding fra klumpen fra DNA som ble brukt til å danne fjæren.. Hvorvidt peri-centromerer er sammensatt av gjentatte sekvenser eller ikke, er et sekundært trekk som skiller seg ut tvers gjennom eukaryotisk fylogeni."
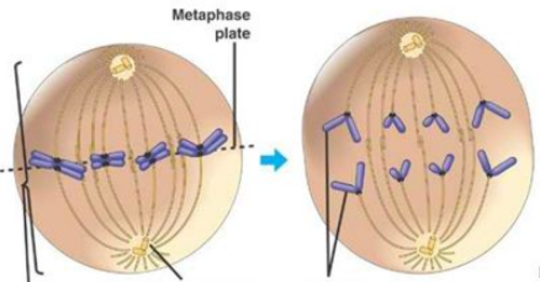 Sammenkveiling
Sammenkveiling
En annen artikkel i Current Biology -lenke diskuterte rollen til centromer sammenkobling i meiose, celledelingsprosessen for seksuell reproduksjon. Evatt et al. fant at "elastiske rep" dannes mellom sammenkoblede kromosomer, og at disse kromatin repene er avhengige av kohesin. Repene lar kromosomer orientere seg ordentlig på spindelen. For mer om rep (tethers) i biokjemi, se denne artikkelen-lenke.
Bilde 3. Celledeling (mitose)
Cohesins arbeid blir aldri gjort
En artikkel i PNAS -lenke fortalte hvordan forberedelse til celledeling er en pågående prosess under interfase - perioden mellom celledelinger. "Aktivitetsdrevet organisasjon" involverer flere trinn, som krever ATP for energi, og inkluderer "komprimering, segregering og undertrykkelse." Det minner meg om hvordan Rose Parade -komiteen begynner å planlegge for neste parade så snart den siste er over. Mer framsyn krevdes for å se behovet for disse prosessene.
Nok en gang spiller Cohesin en viktig rolle gjennom cellesyklusen. "Modellen vår antyder at den raske kinetikken til aktiv sløyfeekspansjon, sammenlignet med den langsomme avslapningen av kromatinløyfer opprettholder en tett kromatin-organisasjon," sier Chan og Rubenstien. "Dette arbeidet presenterer et fysisk rammeverk som forklarer hvordan kohesin bidrar til effektiv transkripsjonell regulering." Se her -lenke for hvordan sløyfeekstrudering er en smart prosess som brukes av ingeniører for å forhindre skade på datamaskinbånd. Cellen bruker denne strategien for å ivareta DNA fra brudd under transkripsjon og replikasjon.
Hold igjen nedtellingen
 Det minner om en nedtellings pause, for endelige kontroller ved en avfyringsrampe, tilsvarende kan pattedyrceller somle i en rolig tilstand, før de vender 'masterbryteren' for å lansere mitose. Forskere ved Weill Cornell Medicine -lenke fant at hovedbryteren E2F, et transkripsjonsfaktorprotein, kan "forbli i en potensielt langvarig tilstand av delvis og reversibel aktivering" før de blir fullt engasjert. Det minner meg om en flykontroller på NASA-utspørring av alle undersystemingeniørene for å rope 'Gå!' før forpliktelsen til å avfyres.
Det minner om en nedtellings pause, for endelige kontroller ved en avfyringsrampe, tilsvarende kan pattedyrceller somle i en rolig tilstand, før de vender 'masterbryteren' for å lansere mitose. Forskere ved Weill Cornell Medicine -lenke fant at hovedbryteren E2F, et transkripsjonsfaktorprotein, kan "forbli i en potensielt langvarig tilstand av delvis og reversibel aktivering" før de blir fullt engasjert. Det minner meg om en flykontroller på NASA-utspørring av alle undersystemingeniørene for å rope 'Gå!' før forpliktelsen til å avfyres.
"Det virker sannsynlig, ifølge forskerne, at denne mellomliggende pregede tilstand gir celler tid til å føle og integrere de vanlige, svingende celledelings-inngangssignalene, jevne ut denne 'støyen' og redusere sjansen for upassende divisjon. Men forskerne mistenker at denne tilstanden også har andre funksjoner, inkludert for å lette DNA-reparasjon, siden celler i denne tilstanden viser tegn til aktiverte DNA-reparasjonsprosesser."
Bilde 4. Illustrasjon av DNA-reparasjon (Credit: Otangelo Grasso)
Oppsummert
Ingen av forskningsartiklene som er sitert i denne artikkelen appelerte til evolusjon for å forklare funnene. Som vanlig er mengden darwinisering omvendt proporsjonal med mengden biologisk detalj i en forskningsartikkel. Intelligent design (framsyn, funksjon og finesse) finnes i detaljene.

Om forfatteren: DAVID COPPEDGE -kredit til David Coppedge (Bilde 5)
David Coppedge er en frilans vitenskapsreporter i Sør-California. Han har vært styremedlem i Illustra Media siden grunnleggelsen og fungerer som deres vitenskapskonsulent. Han jobbet ved NASAs Jet Propulsion Laboratory (JPL) i 14 år, på Cassini-oppdraget til Saturn, til han ble kastet ut i 2011 for å dele materiale om intelligent design, en diskriminerende handling som førte til en nasjonalt publisert rettssak i 2012. Discovery Institute støttet saken hans, men en ensom dommer dømte ham mot ham, uten forklaring. En naturfotograf, friluftsmann og musiker, David har B.S. grader i realfagsutdanning og i fysikk og holder presentasjoner om ID og andre vitenskapelige emner.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund